
Snail sinistrality – levotočivost plžů (Gastropoda)
Díky genetikům je další mysterium v háji…
Směr stáčení ulity určuje formin Lsdia1. Je to ten jako dlouho hledaný gen pro dextrální/sinistrální svinování ulit plžů… doi:10.1242/dev.175976
Sinistralita – levotočivost plžů – je klíčovou složkou plžení jxd. Bilogy: „Většina plžů je stočena ve směru hodinových ručiček, avšak u některých druhů se vyskytují vzácné genetické varianty s obráceným vinutím. Nyní byl identifikován molekulární determinant směru navíjení, cytoskeletální regulátor formin.“ (Maderspacher, 2016). V impaktovaném časopise díky tomu nejde své místo i nálezová zpráva, že v Polsku byl nalezen sinistrální hlemýžď. (Szybial et all. 2015). Chiralita je zkoumána od počátků vzniku malého šneka, už před pářením. Výzkum ukázal, že předkopulační chování plovatky Lymnaea stagnalis je lateralizováno. Asymetrie chování odpovídá sinistrální nebo dextrální chiralitě jedince a je zjevně také řízena lokusem meternálního efektu. To ukazuje, že lateralizované sexuální chování L. stagnalis je od počátku vzniku jedince a je přímým důsledkem asymetrie celého těla. (Davison, et all. 2008)


Výzkum sinistrality pokryl všechny představitelné aspekty malakologického bádání. Pozorování sinistrálního vzorku Arianta arbustorum vědci prokázali, že kopulace zrcadlově převrácených (dextrální x sinistrální) jedinců je velmi komplikovaná. (Gittenberger, Margy, 2011). Pozorování pokusů o kopulaci Arianta arbustorum ukázalo, že jedinci s opačnou chiralitou se nemohou pářit úspěšně. (Margry, Gittenberger 2011). Druhy s plochými ulitami se páří „tváří v tvář“. Tato sexuální symetrie zabraňuje interchirálnímu páření, protože pohlavní orgány vystavené sinistrálem na levé straně nemohou být spojeny s těmi, které jsou vystaveny dextrálníém jedincem na pravé straně. Výhoda proti chirální menšině, která vyplývá z páření, tedy stabilizuje chirální monomorfismus. Druhy s vysokými ulitami jsou nerecipročně spojeny: „samec“ se kopuluje tím, že namontuje plášť „samice“, vzájemně se vyrovnává stejným směrem. Tato sexuální asymetrie umožňuje interchirální kopulaci jen s malými změnami chování. Proto alely reverze přetrvávají déle v populacích druhů s vysokou ulitou. (Asami, Cowie 1998).



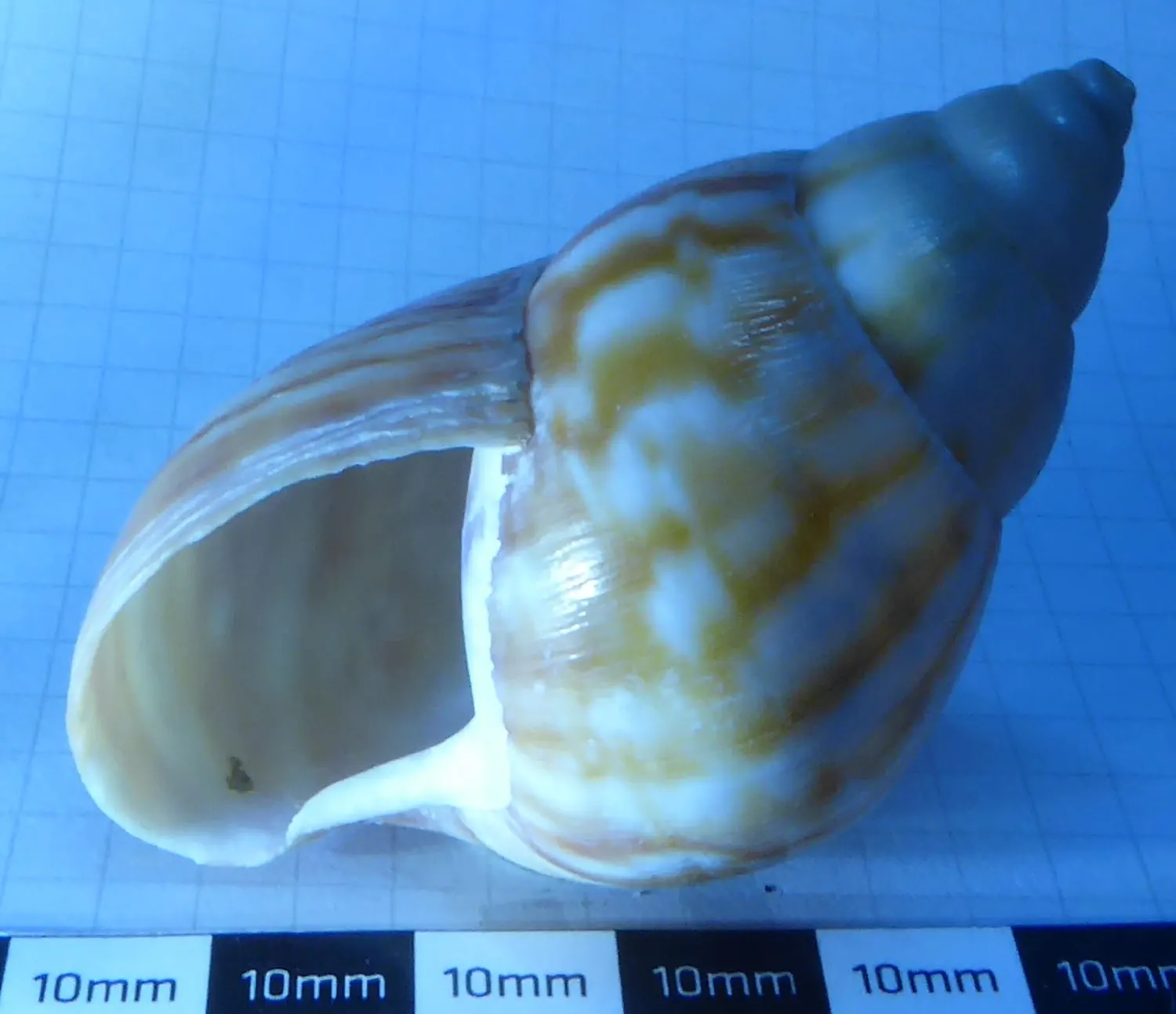
Fosilní sinistrální Conidae – homolice
O sinistralitě se uvažuje i jako o možném zdroji speciace. U několika málo druhů plžů je chiralita určována jediným genetickým lokusem se zpožděnou dědičností, genotyp se projevuje v potomcích matky. Pokusy o izolování lokusů zapojených do chirality plžů se sněží zjistit, jak symetrie vytvořena. Obyčejně se předpokládá, že silná selekce závislá na frekvenci by měla působit proti vytvoření nových chirálních typů, protože chirální menšina má potíže s nalezením vhodného párového partnera (jejich genitálie jsou na „špatné“ straně). Smíšené populace by proto neměly přetrvávat. Ale velmi málo druhů pozemských hlemýždí, zejména subgenus Amphidromus sensu stricto, se nejen zdá, že se náhodně spojí mezi různými chirálními typy, ale mají také stabilní chirální dimorfismus v rámci populace, což naznačuje zapojení vyrovnávacího faktoru. Na druhém konci spektra, u mnoha druhů, se různé chirální typy nedokáží spojit, a tak by se mohly navzájem reprodukovat. Zatímco empirické údaje, modely a simulace naznačují, že se někdy musí objevit chirální obrat, je málo pravděpodobné, že povede k takzvanému „jednogenovéu“ speciaci. Nicméně chirální zvrácení by mohlo být k speciaci (nebo k divergenci po speciaci) přispět. (Davidson, 2005). Jednorázová speciace se považuje za nepravděpodobnou, ale vyskytla se u hlemýžďů, u nichž byl gen pro pravolevou chiralitu obrácen a způsobil vznik několika nových druhů. Tato změna může být usnadněna malou velikostí populace a maternálním efektem – „opožděnou dědičností“, kdy je fenotyp jedince určován genotypem jeho matky. (Yamamichi,Sasaki 2013).



Chiralita může být starým znakem odvozeným z bilaterie. Vypnutí mutace v jedné kopii tandemově duplicitního diafanu – příbuzného forminu je dokonale spojeno se zlomem symetrie ulity plže. To je podpořeno zjištěním, že léčba anti-forminem přeměňuje embrya dextrálního šneku na sinistrální fenokopie a un žab inhibice forminu má chirality-randomizing efekt u raných (pre-cilia) embryí. V rozporu s očekáváními založenými na stávajících modelech jsme objevili asymetrickou expresi genů v embryích s hlemýžďem dvou a čtyř buněk, které předcházejí morfologické asymetrii. Vzhledem k tomu, že vlákno formin-aktinu bylo prokázáno, že je součástí inverzního přepínače asymetrie, jsou společně tyto výsledky v souladu s názorem, že zvířata s různými tělesnými plány mohou odvodit své asymetrie ze stejných intracelulárních chirálních prvků. (Davison, Angus, et all. 2016). Malý výskyt sinistrality v poměru k dextrálitě u plžů je nevysvětleno. U rodu Conus se sinistrální skořápka vyvinula jako převažující charakteristika druhu jen jednou. Fosílie tohoto druhu, Conus adversarius, se nacházejí v horním pliocenu a nejmenších pleistocénových ložiscích v jihovýchodním USA. Conus adversarius měl neplanktonický vývoj larev; to mohlo být kritickým faktorem pro včasnou speciaci, stejně jako sinistrální mořské druhy jiných clade. Většina vzorků aberantně sinistrálních recentních jedinců rodu Conus jsou odvozeny od typicky dextrálních druhů, které mají nonplanktonický vývoj. Pokud byl C. adversarius izolován reproduktivně z dextrálních konspecifických znaků, pak tento druh může poskytnout příklad téměř okamžité sympatrické speciace ve fosilním záznamu. (Hendricks, J. R. 2009).
U druhů plžů, kde jsou sinistralita i dextralita skutečně běžně přítomné, určuje směr navíjení jediný gen s opožděšnou marternální dědičností; neexistuje žádná předvídatelná souvislost mezi vlastním genotypem směru navíjení hlemýždě a jeho skutečným směrem stočení. Kvůli tomuto genetickému oddělení se dá očekávat, že dextrální a sinistrální jedinci budou přesně zrcadlovými obrazy jednoho druhého. Nicméně existují náznaky, že existuje jemný, ale zjistitelný tvarový rozdíl mezi dextrálními a sinistrálními jedinci, které pocházejí ze stejného genofondu.Při zýzkumu 50 dextrálních a 50 sinistrálních jedinců Amphidromus inversus. Když byly ulity podrobeny geometrické morfometrické analýze, sinistrální jedinci skutečně vykazovali mírné, ale významné rozšíření a zkroucení skořápky v blízkosti palatinální a parietální aperturní oblasti. (Schilthuizen, Haase 2010). Řada vědců považuje jednogenovou speciaci za reálnou. U suchozemských hlemýžďů by mohl být zodpovědný za okamžitou speciaci jediný gen pro směrování vinutí vlevo-vpravo. Experimenty ukazují, že sinistrální hlemýždi Camaenidae přežívají při predaci hady Pareas iwasakii (Colubroidea: Pareatidae) lépe než dextrální. (Hoso, et all., 2010). V provedené simulaci se diskutovala speciace na základě jednoho genu chirality. Konkrétně se zkoumalo šest faktorů, a to: (i) absolutní a relativní velikost populace; (ii) úspěch páření, o kterém je známo, že souvisí s tvarem pláště, zejména poměrem výška / šířka; (iii) vliv maternálního efektu, zákládajícím chiralitu; iv) (nízká) pohyblivost šneků; v) rozdíly v způsobilosti (heterosa); a (vi) zda invazivní mutantní alela je dominantní nebo recesivní. Dopad těchto faktorů byl kvantifikován.

Malá populace s nepatrným počtem predátorů a dominantní postavení alely mutantní chirality mají zásadní význam pro příležitostnou fixaci druhu. (van Batenburg, Gittenberger, 1996). Podle jiných autorů jsou však důkazy na podporu jednogenového speciace řídké, většinou založené na monogenních mitochondriálních studiích a vzorcích chirálních variací mezi jednotlivými druhy.
Studie použila teoretický model, který ukázal, že chirální fenotyp potomstva je určován maternálním genotypem,. Mohou se tedy objevit příležitostné chirální obraty, a umožní se tok genů mezi morfovými zrcadlovými obrazy, které brání speciaci. Empiricky se ukázalo, že mezi různými chirálními typy japonských druhů Euhadra existuje nedávný nebo probíhající genový tok. Zaznamenány byly také důkazy o spojení mezi zrcadlovými morfy, které přímo ukazují potenciál pro tok genů. Takže teoretické modely naznačují tok genů mezi opačně navinutými hlemýžďmi a empirická studie ukázaly, že se mohou spojit a že u rodu Euhadra dochází k toku genů mezi jedinci s opačným vinutím. Je zapotřebí víc než jediný gen, než může být chirální změna směru vinutí ulity považována za vytvoření nového druhu. (Richards, et all. 2018).

Typy sinistrality
- Sinistrální jedinec jako monstrum simplex
Někteří plži jsou nativně, „od přírody“, výhradně dextrální, mají výhradně dextrální – pravotočivou – ulitu. Jde například o známého hlemýždě zahradního (Helix pomatia). U těchto druhů se však výjimečně vyskytne jedinec, který má ulitu levotočivou. Jde v řadě případů (ne však vždy) o mutanta, o monstrum simplex v plném slova smyslu, ovšem plně viabilní. Přestože malakologie již dlouho říká, že tato mutace není gametická, a že sinistrální jedinci sinistralitu nepřenesou do F1 na své potomky,dochází vzhledem k celkové atraktivitě fenomenu k opakovaným pokusům o odchov sinistrálních Helix s nadějí, že i F1 bude sinistrální. I odchovy jxd ze sinistrálních rodičů Helix aspersa byly pravotočiví – normální – šneci…



- Sinistralita jako řídká divoká alela
To jest málo hojná sinistrální část populace, nikoliv mutant. Sinistralita jako řídká divoká alela se vyskytuje u řady rodů – například Phylla, Lymnaea a dalších. U těchto rodů se dle literatury má vyskytovat v běžně dextrální populaci sinistralita s frekvencí cca 2 procenta.



- Sinistralita jako dominantní znak populace
Sinistralita jako dominantní znak populace existuje u (dle docenta Davisona) cca 10 procent druhů plžů, ti jsou „od přírody“ levotočiví. V takových taxonech je monstrum simplex jedinec dextrální. Jde například o čeleď závornatkovitých (Clausiliidae). 26 (!) druhů žijících v ČR je nativně sinistrálních, sinistrální druhy převažují globálně. Dále jsou nativně sinistrální dva druhy rodu vrkoč (Vertigo angustior a pusilla).

Reference:
Asami, Takahiro, Cowie, Robert H. (1998).Evolution of Mirror Images by Sexually Asymmetric Mating Behavior in Hermaphroditic Snails. The American Naturalist 1998. DOI: 10.1086/286163
Blum, Martin., Vick, Phillip (2015). Left–Right Asymmetry: Cilia and Calcium Revisited. Journal for Current Biology. Vol. 25, Iss.5, 2015, pp. R205-R207. https://doi.org/10.1016/j.cub.2015.01.031
Čevela, Vlastimil (2018). Osobní sdělení majitele zemědělské farmy Farma Nahošovice zabývající se velkochovem kaviárových hlemýžďů Helix aspersa. (Farma Nahošovice, Nahošovice č.p. 25, Dřevohostice, Czech Republic) dne 11. prosince 2018.
Davison, Angus, Schilthuizen, Menno, (2005). The convoluted evolution of snail chirality. The Science of Nature. DOI: 10.1007/s00114-05-0045-2
Davison, Angus, Frend, Hayley T., Moray, Camile, Wheatley, Hannah, Searle, Laura J. and Eichhorn, Markus P. (2008): Mating behaviour in Lymnaea stagnalis pond snails is a maternally inherited, lateralized trait. Biology Letters: Animal behaviour. http://dx.doi.org/10.1098/rsbl.2008.0528
Davison, Angus., McDowell,Gary S., Holden, Jennifer M., Johnson, Harriet F., Koutsovoulos, Georgios D., Liu, M. Maureen., Hulpiau, Paco., Van Roy, Frans., Wade, Christopher M., Banerjee, Ruby., Yang, Fengtang., Chiba, Satoshi., Davey,John W., Jackson, Daniel J., Levin, Michael and Blaxter, Mark L.(2016). Formin Is Associated with Left-Right Asymmetry in the Pond Snail and the Frog. Current Biology. Vol. 26, Iss. 5, pp. 654-660, MARCH 07, 2016.Online: https://pubmed.ncbi.nlm.nih.gov/26923788/
Dietl, Gregory P,. Hendricks, Jonathan R. (2006). Crab scars reveal survival advantage of left-handed snails. Biol Lett. 2006 Sep 22; 2(3): 439–442. doi: 10.1098/rsbl.2006.0465
Gittenberger, Edmund (1988). Notes and commnets sympatric speciation in snails; a largely neglected model. Evolution, 42(4), 1988, pp. 826-828
Gittenberger, E., Margy, C.J.P.J.(2011). Premating isolation reconfirmed in Arianta arbustorum (Linnaeus, 1758) (Gastropoda, Pulmonata, Helicidae). Basteria, vol. 75 (4-6), 2011, 57-58. https://www.researchgate.net/publication/317239100_Premating_isolation_reconfirmed_in_Arianta_arbustorum_Linnaeus_1758_Gastropoda_Pulmonata_Helicidae
Haase, Martin, Baur, Bruno (1995). Variation in spermathecal morphology and storage of spermatozoa in the simultaneously hermaphroditic land snail Arianta arbustorum (Gastropoda: Pulmonata: Stylommatophora). Invertebrate Reproduction & Development 33.Vol. 28. Iss. 1. DOI: 10.1080/07924259.1995.9672461
Hendricks, J. R. (2009), Sinistral snail shells in the sea: developmental causes and consequences. Lethaia. Vol. 42. pp. 55-66. doi:10.1111/j.1502-3931.2008.00103.x
Hoso,Masaki., Kameda,Yuichi,. Wu,Shu-Ping., Asami,Takahiro., Kato,Makoto, and Hori, Michio (2010).A speciation gene for left–right reversal in snails results in anti-predator adaptation. Nature Communications. 2010. Vol. 1. pp. 133. doi:10.1038/ncomms1133
Maderspacher, Florian (2016). Snail Chirality: The Unwinding. Current Biology Dispatches https://www.cell.com/current-biology/pdf/S0960-9822(16)30060-4.pdf
Margry, C.J.P.J., Gittenberger E.(2011). Premating isolation reconfirmed in Arianta arbustorum (Linnaeus, 1758) (Gastropoda, Pulmonata, Helicidae). Basteria. Vol. 75 (4-6).
Naganathan, Sundar Ram, Middelkoop,Teije C., Fürthauer, Sebastian, Grill, Stephan W. (2016).Actomyosin-driven left-right asymmetry: from molecular torques to chiral self organization. Current Opinion in Cell Biology. https://doi.org/10.1016/j.ceb.2016.01.004
Nakadera, Yumi, Blom, Christiaan, Koene, Joris M. (2014). Duration of sperm storage in the simultaneous hermaphrodite Lymnaea stagnalis Journal of Molluscan Studies, Volume 80, Issue 1, pp. 1–7. https://doi.org/10.1093/mollus/eyt049
Richards,Paul M., Morii,Yuta., Kimura,Kazuki., Hirano,Takahiro., Chiba,Satoshi and Davison, Angus (2018). Single-gene speciation: Mating and gene flow between mirror-image snails. Evolution Letterrs, Volume 2, Issue 1, Pages: 1-48, February 2018
Schilthuizen, M., Haase, M. (2010). Disentangling true shape differences and experimenter bias: are dextral and sinistral snail shells exact mirror images? Journal Zool.Vol. 282(3). pp. 191–200. doi: 10.1111/j.1469-7998.2010.00729.x
Schilthuizen, Menno., van Heuven, Bertie-Joan (2011). Dextral and sinistral Amphidromus inversus (Gastropoda: Pulmonata: Camaenidae) produce dextral sperm. Zoomorphology. Vol. 130, p. 283. https://doi.org/10.1007/s00435-011-0140-1
Szybiak, Krystyna, Błoszyk, Jerzy, Kalinowski, Tomasz, Książkiewicz, Zofia. (2015). New data on sinistral and scalariform shells among roman snail Helix pomatia Linnaeus, 1758 in Poland. Folia Malacologica Vol. 23: pp. 47-50. http://dx.doi.org/10.12657/folmal.023.004
Takashi Okumura,Takashi., Utsuno, Hiroki., Kuroda,Junpedi., Gittenberger, Edmund., Asami, Takahiro and Matsuno, Kenji(2008). The Development and Evolution of Left-Right Asymmetry in Invertebrates: Lessons From Drosophila and Snails. Developmental Dynamics, Vol. 237, pp. 3497–3515.
van Batenburg, F. H. D., Gittenberger, E. (1996). Ease of fixation of a change in coiling: computer experiments on chirality in snails. Heredity. 1996. Vol. 76, pp. 278. The Genetical Society of Great Britain. http://dx.doi.org/10.1038/hdy.1996.41
Vermeij, Geerat J. (1975).Evolution and distribution of left-handed and planispiral coiling in snails. Nature. Nature Publishing Group. http://dx.doi.org/10.1038/254419a0
Wandelt, J., Nagy, L.M. (2004). Left-Right Asymmetry: More Than One Way to Coil a Shell. Current Biology. Volume 14, Issue 16, 2004, Pp. R654-R656. https://doi.org/10.1016/j.cub.2004.08.010
Whelan, Nathan V., Strong, Ellen E. (2014). Seasonal reproductive anatomy and sperm storage in pleurocerid gastropods(Cerithioidea: Pleuroceridae). Can. J. Zool. 92: 989–995 (2014) dx.doi.org/10.1139/cjz-2014-0165
Yamamichi, Masato., Sasaki, Akira (2013). SINGLE-GENE SPECIATION WITH PLEIOTROPY: EFFECTS OF ALLELE DOMINANCE, POPULATION SIZE, AND DELAYED INHERITANCE. Evolution. The Society for the Study of Evolution. Evolution 67-7: 2011–2023. https://onlinelibrary.wiley.com/doi/full/10.1111/evo.12068